Rows: 333
Columns: 8
$ species <fct> Adelie, Adelie, Adelie, Adelie, Adelie, Adelie, Adel…
$ island <fct> Torgersen, Torgersen, Torgersen, Torgersen, Torgerse…
$ bill_length_mm <dbl> 39.1, 39.5, 40.3, 36.7, 39.3, 38.9, 39.2, 41.1, 38.6…
$ bill_depth_mm <dbl> 18.7, 17.4, 18.0, 19.3, 20.6, 17.8, 19.6, 17.6, 21.2…
$ flipper_length_mm <int> 181, 186, 195, 193, 190, 181, 195, 182, 191, 198, 18…
$ body_mass_g <int> 3750, 3800, 3250, 3450, 3650, 3625, 4675, 3200, 3800…
$ sex <fct> male, female, female, female, male, female, male, fe…
$ year <int> 2007, 2007, 2007, 2007, 2007, 2007, 2007, 2007, 2007…25 Supervised Learning
Imagine you have measured a few hundred penguins on a remote island — the length and depth of each bird’s bill, the length of its flipper. Now a new bird shows up. Can you predict its body mass from those measurements alone? Or guess which species it belongs to? That is exactly the kind of question supervised learning answers: you have examples where you know the answer, and you want a rule that predicts the answer for a new case you haven’t seen.
The same shape of problem shows up everywhere in biology. Predict a tumour’s response to a drug from its gene-expression profile. Predict a patient’s blood pressure from a panel of clinical measurements. Classify a cell as healthy or diseased from its morphology. In every case you have features (the inputs you measure) and a target (the answer you want to predict), and a pile of past examples to learn from.
In the previous chapter we met the three paradigms of machine learning and the all-important habit of splitting data to avoid fooling ourselves. This chapter is a guided tour of the workhorse supervised algorithms — linear regression, k-nearest neighbours, ridge/LASSO/elastic-net, decision trees, and random forests. For each one you’ll get a plain-language intuition and a small, runnable example on a single shared dataset, so you can see the algorithms as variations on one theme rather than a zoo of unrelated tricks. (The later mlr3verse chapter shows how to run all of these through one tidy interface; here we stay close to base R so the moving parts stay visible.)
25.1 What you’ll learn
- Describe any supervised algorithm in terms of its hypothesis space, objective function, and learning algorithm.
- Fit and interpret a linear regression with
lm(). - Make predictions with k-nearest neighbours using
class::knn(), and explain why feature scaling matters. - Fit ridge, LASSO, and elastic-net models with
glmnet(), and read off which features each one keeps. - Grow and read a decision tree with
rpart, and a random forest withranger.
25.2 A framework for understanding supervised algorithms
Every supervised algorithm in this chapter — and almost every one you’ll meet elsewhere — can be taken apart into the same three pieces. Once you can name those three pieces for an algorithm, you understand most of what makes it tick, and you can compare any two algorithms cleanly.
ImportantThe three pillars of a supervised algorithm
Hypothesis space (the model family). The set of all candidate models the algorithm is allowed to consider. For linear regression it’s “all straight lines”; for a decision tree it’s “all branching yes/no rule sets.” This choice bakes in an assumption — a bias — about what the answer can look like.
Objective function (the goal). A number that says how good a candidate model is on the training data — usually how badly it errs, so smaller is better. For regression that’s often the mean squared error. Learning means finding the model that makes this number as small as possible.
Learning algorithm (the optimiser). The actual procedure that searches the hypothesis space for the model with the best objective value. Sometimes it’s a one-shot formula (ordinary linear regression); sometimes it’s a long iterative grind (neural networks).
Here’s the payoff of this framing. Ridge and LASSO regression, which we’ll meet shortly, use the same hypothesis space and same style of optimiser as plain linear regression. The only thing they change is the objective function — they add a penalty for being too complicated. That one small change is what lets them avoid overfitting. Keep the three pillars in mind and each new algorithm becomes a small variation, not a whole new world.
25.4 Linear regression
Plain-language intuition. Linear regression draws the best straight-line relationship between your features and a continuous target. “Best” means the line that makes the total squared prediction error as small as possible. Each feature gets a coefficient — a slope — telling you how much the target changes when that feature goes up by one unit. It is the simplest, most interpretable model in the toolbox, and a sensible first thing to try.
In the language of the three pillars: the hypothesis space is “all straight-line combinations of the features,” the objective function is the sum of squared errors (Equation 25.1), and the learning algorithm is a tidy formula that R solves instantly.
\[L = \sum_{i=1}^{n}{(\hat{y}_i - y_i)}^2 \tag{25.1}\]
Here \(\hat{y}_i\) is the predicted value, \(y_i\) is the true value, and \(n\) is the number of observations. Let’s predict penguin body mass from flipper length and bill dimensions:
(Intercept) flipper_length_mm bill_length_mm bill_depth_mm
-6359.49030 50.52663 1.88153 18.08102 Read the coefficients as “holding the others fixed, one extra millimetre of flipper length is associated with about 51 grams more body mass.” That is the chief virtue of linear regression: the model is its explanation.
Now the honest test — how far off are the predictions on the held-out birds? We summarise that with the root mean squared error (RMSE), roughly the typical prediction error in grams:
[1] 387.8589So the model’s predictions are typically off by around that many grams — for birds weighing 3,000–6,000 g, not bad for three measurements and a straight line.
25.5 K-nearest neighbours
Plain-language intuition. k-nearest neighbours (k-NN) is the “you are who your neighbours are” algorithm. To predict the answer for a new penguin, it finds the k most similar penguins in the training data — the ones closest to it in measurement space — and lets them vote. For classification, the new bird is assigned the most common species among its k neighbours. For regression, it gets the average of their values.
k-NN is unusual: it has no real “training” step and no equation to fit. It just stores the data and measures distances at prediction time. Its one knob is k: a small k (say 1) follows every wiggle in the data and can overfit; a large k smooths things out and can underfit. k is a hyperparameter you tune, exactly like we discussed in the previous chapter.
WarningScale your features first
k-NN decides “closeness” using distance, so a feature measured in large numbers will dominate one measured in small numbers — purely because of its units, not its importance. Flipper length (≈ 200 mm) would swamp bill depth (≈ 17 mm) if we left them as-is. The fix is to standardise every feature to a common scale before computing distances. Forgetting this is the single most common k-NN mistake.
Let’s classify penguin species from the three measurements. We standardise using the training set’s mean and standard deviation, then apply the same shift and scale to the test set:
library(class)
features <- c("flipper_length_mm", "bill_length_mm", "bill_depth_mm")
train_means <- colMeans(train[features])
train_sds <- apply(train[features], 2, sd)
scale_with <- function(df) scale(df[features], center = train_means, scale = train_sds)
train_x <- scale_with(train)
test_x <- scale_with(test)
set.seed(42)
knn_pred <- knn(train = train_x, test = test_x, cl = train$species, k = 5)
mean(knn_pred == test$species) # accuracy: fraction classified correctly[1] 0.98With k = 5 neighbours, k-NN gets the species right for the large majority of held-out birds. The three penguin species really do cluster into distinct regions of measurement space, which is exactly the situation where “ask your neighbours” works beautifully.
25.6 Penalized regression
Plain linear regression has a weakness: when you have many features — especially more features than observations, the common situation in genomics — it can chase noise and overfit. Penalized regression fixes this by tweaking the objective function (pillar two). It keeps the usual squared-error term but bolts on a penalty that grows as the coefficients get large, so the fitting procedure now has to balance two goals at once: fit the data well and keep the coefficients small. The net effect is a model that is pressured to stay simple.
A single dial, lambda (\(\lambda\)), sets how hard that pressure pushes. When \(\lambda = 0\) the penalty vanishes and you are back to ordinary linear regression; as \(\lambda\) grows, the coefficients are squeezed ever harder toward zero. The three methods below — ridge, LASSO, and elastic net — share this idea and differ only in the shape of the penalty they add.
We’ll fit all three with the glmnet package. glmnet does not take a formula; it wants a numeric matrix of features x and a response vector y. The model.matrix() step below builds that matrix (and drops the intercept column with [, -1]):
library(glmnet)
x_train <- model.matrix(body_mass_g ~ flipper_length_mm + bill_length_mm +
bill_depth_mm, data = train)[, -1]
y_train <- train$body_mass_g
x_test <- model.matrix(body_mass_g ~ flipper_length_mm + bill_length_mm +
bill_depth_mm, data = test)[, -1]
y_test <- test$body_mass_g25.6.1 Ridge regression
Plain-language intuition. Ridge uses an L2 penalty: the sum of the squared coefficients (Equation 25.2). Because squaring punishes large coefficients much more than small ones, ridge pulls every coefficient toward zero but never lets any of them land exactly on zero. Every feature therefore survives — ridge simply turns down their volume rather than dropping any. That makes it a good fit when you suspect many features each carry a little of the signal.
\[L = \sum_{i=1}^{n}{(\hat{y}_i - y_i)}^2 + \lambda\sum_{j=1}^{k} \beta_j^2 \tag{25.2}\]
In glmnet, ridge is alpha = 0. We use cv.glmnet() to let cross-validation pick a good \(\lambda\) automatically:
4 x 1 sparse Matrix of class "dgCMatrix"
lambda.min
(Intercept) -4592.10682
flipper_length_mm 41.68954
bill_length_mm 13.28295
bill_depth_mm -10.47499Notice that all three coefficients are present and non-zero — ridge shrank them but kept them. Its test error:
pred_ridge <- predict(fit_ridge, newx = x_test, s = "lambda.min")
rmse(y_test, pred_ridge)[1] 396.011625.6.2 LASSO regression
Plain-language intuition. LASSO (Least Absolute Shrinkage and Selection Operator) swaps in an L1 penalty: the sum of the absolute coefficients (Equation 25.3). Switching from squares to absolute values sounds like a minor change, but it has a dramatic consequence — the L1 penalty can drive a coefficient all the way to zero, which is the same as striking that feature out of the model entirely. LASSO therefore performs automatic feature selection, leaving you with a leaner model built only from the features that pull their weight.
\[L = \sum_{i=1}^{n}{(\hat{y}_i - y_i)}^2 + \lambda\sum_{j=1}^{k} |\beta_j| \tag{25.3}\]
LASSO is alpha = 1 in glmnet:
4 x 1 sparse Matrix of class "dgCMatrix"
lambda.min
(Intercept) -5499.365152
flipper_length_mm 47.897811
bill_length_mm 1.414023
bill_depth_mm . Any coefficient shown as . has been zeroed out and dropped. With only three strong features here, LASSO may keep all of them — but on a genomics dataset with thousands of features, this is how you go from “20,000 genes” to “the handful that actually predict the outcome.”
TipRidge or LASSO? Let the data decide
Neither always wins. LASSO shines when a few features carry the signal and the rest are noise (so zeroing them helps). Ridge shines when many features each contribute a bit. When you’re unsure — which is most of the time — fit both with cross-validation and compare their test error, exactly as we’re doing here.
25.6.3 Elastic net
Plain-language intuition. Why choose? Elastic net blends the L1 and L2 penalties, so it can zero out useless features (the LASSO trick) while gracefully handling groups of correlated features (the ridge strength). When two features are highly correlated, pure LASSO tends to arbitrarily keep one and drop the other; elastic net is happier keeping both, which is common and useful in biology where genes in the same pathway move together.
The mixing dial is alpha, between 0 and 1: alpha = 0 is pure ridge, alpha = 1 is pure LASSO, and anything in between is elastic net. Let’s use a 50/50 blend, alpha = 0.5:
4 x 1 sparse Matrix of class "dgCMatrix"
lambda.min
(Intercept) -6166.200623
flipper_length_mm 49.790369
bill_length_mm 2.389242
bill_depth_mm 14.153060pred_enet <- predict(fit_enet, newx = x_test, s = "lambda.min")
rmse(y_test, pred_enet)[1] 388.1342Elastic net gives you a coefficient profile somewhere between ridge’s “keep everything, shrunk” and LASSO’s “keep a few.” On this small three-feature problem all four regression models land in roughly the same place; the penalized methods earn their keep when features are many and correlated.
25.7 Classification and regression trees (CART)
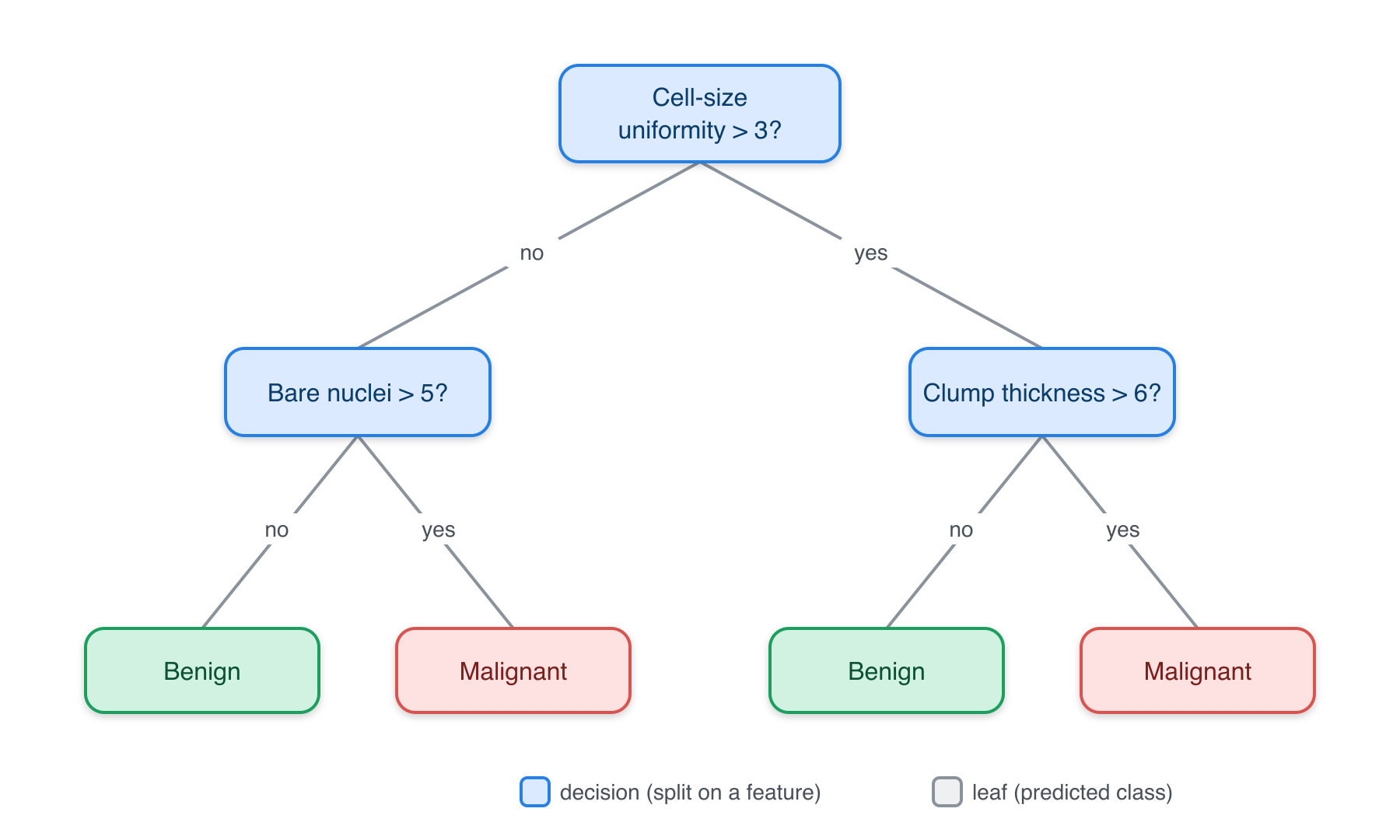
Plain-language intuition. A decision tree predicts by asking a series of simple yes/no questions about the features, like a flow chart or the game of Twenty Questions. “Is the flipper longer than 206 mm? If yes, is the bill deeper than 17 mm?” Each question splits the data into purer and purer groups, until a final leaf gives the prediction — the majority species (for classification) or the average value (for regression).
In three-pillar terms: the hypothesis space is “all such question trees,” the objective at each split is to make the resulting groups as pure as possible (measured by the Gini index or entropy), and the learning algorithm greedily picks the best split at each step. Trees are prized because you can read them — they match how people naturally reason.
We’ll grow a classification tree for species with rpart (which ships with R):
[1] 0.94The tree learned a short list of measurement thresholds that separate the species, and it classifies the held-out birds with high accuracy. We can even print the learned rules:
fit_treen= 233
node), split, n, loss, yval, (yprob)
* denotes terminal node
1) root 233 131 Adelie (0.43776824 0.19742489 0.36480687)
2) flipper_length_mm< 206.5 142 42 Adelie (0.70422535 0.29577465 0.00000000)
4) bill_length_mm< 44.2 101 3 Adelie (0.97029703 0.02970297 0.00000000) *
5) bill_length_mm>=44.2 41 2 Chinstrap (0.04878049 0.95121951 0.00000000) *
3) flipper_length_mm>=206.5 91 6 Gentoo (0.02197802 0.04395604 0.93406593)
6) bill_depth_mm>=17.2 7 3 Chinstrap (0.28571429 0.57142857 0.14285714) *
7) bill_depth_mm< 17.2 84 0 Gentoo (0.00000000 0.00000000 1.00000000) *Figure 25.1 shows the same idea drawn out for a clinical example: each internal node tests one feature, and following the branches leads to a leaf with a predicted class.
WarningA single tree is twitchy
Decision trees have a real flaw: they’re unstable. Change a few training rows and you can get a noticeably different tree. A single deep tree also overfits easily. The fix — averaging many trees together — is exactly the idea behind random forests, next.
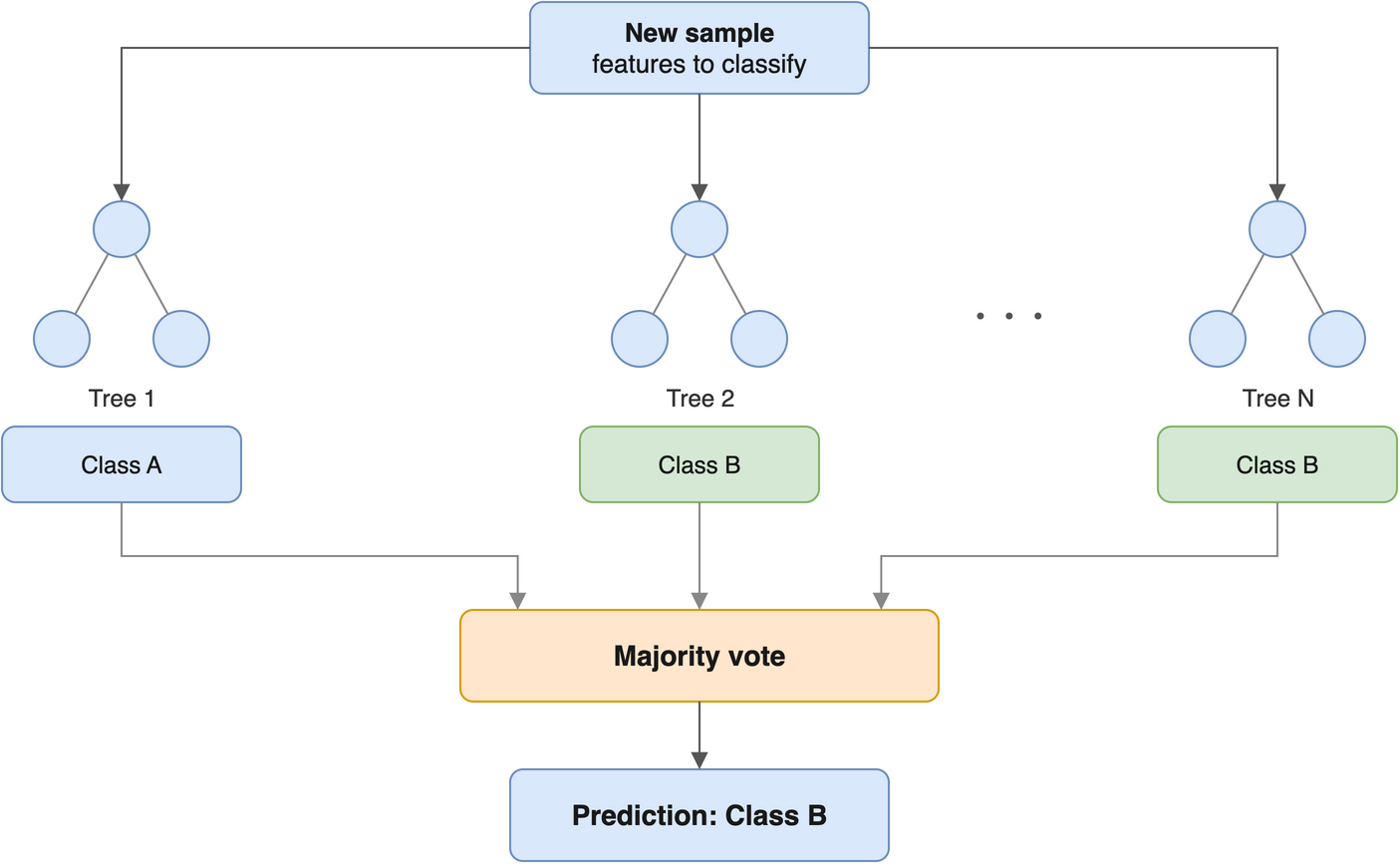
25.8 Random forests
Plain-language intuition. If one decision tree is twitchy, build hundreds of them and let them vote. A random forest grows many trees, each on a random bootstrap sample of the data and each allowed to consider only a random subset of features at every split. That deliberate randomness makes the individual trees disagree in their mistakes — and when you average many imperfect-but-different predictions, the errors cancel out. The result is far more stable and accurate than any single tree, while needing very little tuning. Figure 25.2 sketches the idea: many trees, one combined vote.
We’ll use the fast ranger package. Random forests are randomised, so we set a seed for a reproducible result:
[1] 0.97The forest typically matches or beats the single tree on the held-out birds, with no manual tuning — which is exactly why random forests are such a popular, reliable default for tabular biological data.
25.9 Summary
You now have a working map of the supervised-learning landscape, and you’ve run every algorithm on it with your own hands:
- Every algorithm is a hypothesis space + an objective function + a learning algorithm. Name those three and you understand it.
-
Linear regression (
lm()) fits an interpretable straight-line model; coefficients tell you how each feature moves the target. -
k-nearest neighbours (
class::knn()) predicts from the most similar training examples — remember to scale your features first. -
Penalized regression (
glmnet()) adds a complexity penalty to fight overfitting: ridge (alpha = 0) shrinks all coefficients, LASSO (alpha = 1) zeroes some out for feature selection, and elastic net (0 < alpha < 1) blends the two. -
Decision trees (
rpart) are readable flow charts but unstable on their own; random forests (ranger) average many trees into a stable, accurate, low-fuss predictor.
In every case we trained on one split and judged on a held-out split — the habit that keeps you honest. The next chapters show how to run all of these through the unified mlr3verse interface, so swapping one algorithm for another becomes a one-line change.
25.10 Exercises
-
A simpler linear model. Refit the linear regression for
body_mass_gusing onlyflipper_length_mm. Compute its test RMSE and compare it to the three-feature model. Did dropping two features hurt much?NoteSolution[1] 389.7472Flipper length alone is a strong predictor of body mass, so the simpler model is usually only a little worse. That’s a useful reminder: more features are not automatically better.
-
Tuning k in k-NN. Try
k = 1,k = 5, andk = 25for the species classifier. Which gives the best test accuracy? What goes wrong at the extremes?NoteSolutionk = 1 accuracy = 0.98 k = 5 accuracy = 0.98 k = 25 accuracy = 0.96Very small
k(like 1) follows noise and can overfit; very largekblurs the class boundaries and underfits. A middle value usually wins — which is whykis a hyperparameter worth tuning. -
Watch LASSO select. Push the LASSO penalty up by using
lambda.1se(a more aggressive, simpler model) instead oflambda.min, and look at the coefficients. Are any features dropped?NoteSolutioncoef(fit_lasso, s = "lambda.1se")4 x 1 sparse Matrix of class "dgCMatrix" lambda.1se (Intercept) -4070.4745 flipper_length_mm 41.1108 bill_length_mm . bill_depth_mm .lambda.1seis the largest penalty whose error is still within one standard error of the best — a deliberately simpler model. Any coefficient printed as.has been set to exactly zero and removed. With strong features here it may keep them all, but the mechanism is the same one that prunes thousands of genes down to a few. -
Feature importance from the forest. Refit the random forest with
importance = "impurity"and printfit_rf$variable.importance. Which measurement matters most for telling the species apart?NoteSolutionbill_length_mm flipper_length_mm bill_depth_mm 58.71576 47.88220 40.17413The feature with the largest importance score contributed most to splitting the species apart across the forest’s trees. Importance scores are one of the friendliest things random forests give you — a ranked list of which measurements carry the signal.
-
Regression, three ways. You have test RMSE for linear regression, ridge, and elastic net. Compute it for LASSO too, then state which model you’d ship and why.
NoteSolutionlinear ridge lasso enet 387.8589 396.0116 390.8471 388.1342On this small, low-dimensional problem the four are nearly tied, so you’d pick the simplest and most interpretable — plain linear regression. The penalized methods pull ahead only when features are numerous and correlated, as in real genomic data.